

Introduction
It is now almost100 years ago that the first victim with severe dementia associated with neuronal cell death was studied by Alois Alzheimer. By that time such cases were very unusual and Alois Alzheimer had the chance to see only two patients throughout his life. Now Alzheimer�s disease (AD) is a very frequent disorder, which affects millions worldwide. Only within Germany, about 1.2 million patients suffer from this devastating disorder. During the last decades a sharp increase of affected persons is observed, which still continues and, according to recent statistical predictions, becomes very dramatic in the near future. This is due to the enormous increase in our life expectancy. Work on the molecular mechanisms of AD from my laboratory and many others now strongly indicate that AD is an age dependent syndrome, which will affect almost all of us as long as we life long enough.
Results/Project Status
Previous work of the laboratory
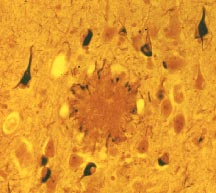
In my laboratory we focus on the generation of a deadly peptide, the Amyloid ß-peptide (Aß), which accumulates during aging in our brains, and becomes deposited as insoluble aggregates called amyloid plaques (Fig. 1) (Haass, 2004). Aggregating Aß is known to be neurotoxic and similar processes of protein aggregation and selective cell death are involved in other neurodegenerative disorders (Aguzzi and Haass, 2003).
Back in 1992, I made the very surprising observation that Aß is produced constantly throughout life (Haass et al., 1992). Moreover, highest levels of Aß generation were observed in human neurons, which constantly secrete Aß by a physiologically totally normal pathway (Haass et al., 1992). These observations changed the common concept of AD, and it is now clear that AD constitutes a major part of normal aging (Haass, 2004).
Since Aß has a central role in AD pathology, we subsequently investigated the mechanisms behind its generation. It quickly became clear that Aß is generated by proteolytic processing involving two types of proteases, ß-, and gamma-secretase (Haass and Selkoe, 1993). Both secretases are major drug targets, since their inhibition should slow down the age related neuropathology caused by aggregating Aß. Our work on gamma-secretase focuses on its identification, function, assembly, and reconstitution. It turned out that gamma-secretase cleaves its substrates twice within the membrane (Okochi et al., 2002), a proteolytic processing pathway so far believed to be biochemically impossible. Moreover, by identifying the active sites of the gamma-secretase activity in presenilins (PS1 and PS2), we were able to define a completely novel class of aspartyl proteases of the GxGD type (Steiner and Haass, 2000; Steiner et al., 2000). By using C. elegans and zebrafish as a model system in combination with biochemical and cell biological technologies, we could demonstrate that presenilins are directly involved in the Notch signaling pathway (Capell et al., 2000; Geling et al., 2002). However, PS alone could not perform proteolysis and we could demonstrate that a complex composed of four different proteins is required to reconstitute gamma-secretase activity in yeast (Edbauer et al., 2003). Currently we are investigating the assembly of the gamma-secretase complex and the interaction sites of the individual components (Kaether et al., 2004; Kaether et al., 2002). In addition we are searching for mechanisms, which could affect the precision of the gamma-secretase cleavage during aging and thus lead to enhanced aggregation and deposition of Aß.
In parallel we are studying the function and regulation of ß-secretase. Here we are specifically interested in regulative mechanisms, which may be responsible for the increased ß-secretase activity during aging. First evidence suggests a posttranscriptional mechanism via the 5� untranslated region of the ß-secretase mRNA (Lammich et al., 2004). The function of ß-secretase and its homologues is investigated in zebrafish, where we knocked down the expression of the corresponding genes and observed very promising phenotypes.
Zebrafish is also used to generate a vertebrate model system for the investigation of age related Aß neurotoxicity. Based on such a model we currently prepare a mutagenesis screen to identify enhancers and inhibitors of Aß mediated cell death in vivo.
Project Status
A: Identification if six different gamma-secretase complexes
gamma-Secretase catalyzes the final endoproteolytic cleavage of the ß-amyloid precursor protein and liberates the neurotoxic amyloid ß-peptide, which accumulates in the brain of patients suffering from AD. Genetic and biochemical studies have demonstrated that gamma-secretase is a protein complex composed of the catalytic subunit presenilin (PS) and three additional subunits, nicastrin (NCT), APH-1 and PEN-2. In mammals, two PS homologues, PS1 and PS2 exist which are part of two distinct gamma-secretase complexes. Likewise two APH-1 homologues (APH-1a and APH-1b) have been identified, with APH-1a existing in two different splice forms (APH-1aL and APH-1aS). So far it was unclear if APH-1 proteins are part of identical or separate gamma-secretase complexes. We could recently demonstrate that APH-1aL, APH-1aS and APH-1b form separate, proteolytically active gamma-secretase complexes containing either one of the two PSs (Shirotani et al., 2004). We could further demonstrate that APH-1aS is the principal APH-1a splice form expressed in human embryonic kidney 293 cells, whereas APH-1aL was identified as the principal splice form in mouse cortex. Surprisingly, we observed a complex pattern of regulation of APH-1a and APH-1b expression. Knockdown of NCT expression caused a decrease in APH-1aS/L levels, whereas APH-1b levels remained unchanged. Knockdown of PEN-2 or a PS1/2 double knockout caused a decrease in APH-1aS/L and an increase of APH-1b levels. Taken together, our results show that dependent on the tissue expression of the individual subunits at least up to six distinct gamma-secretase complexes composed of the known subunits can exist. APH-1b, like PS2 may have evolved as a backup for APH-1a variants. We are now investigating the individual function of these complexes.
B: Novel GxGD Presenilin-like proteases
Signal peptide peptidase (SPP) is an unusual aspartyl protease, which mediates clearance of signal peptides by proteolysis within the endoplasmic reticulum (ER). Like presenilins, which provide the proteolytically active subunit of the gamma-secretase complex, SPP contains a critical GxGD motif in its C-terminal catalytic center. While SPP is known to be an aspartyl protease of the GxGD type, several presenilin homologues / SPP-like proteins (PSHs/SPPL) of unknown function have been identified by database searches. We now investigated the subcellular localization and a putative proteolytic activity of PSHs/SPPLs in cultured cells and in an in vivo model. We demonstrate that SPPL2b is targeted through the secretory pathway to endosomes/lysosomes while SPP and SPPL3 are restricted to the ER. As suggested by the differential subcellular localization of SPPL2b compared to SPP and SPPL3, we found distinct phenotypes upon antisense gripNA mediated knockdown in zebrafish. spp and sppl3 knockdowns in zebrafish result in cell death within the central nervous system, whereas reduction of sppl2b expression causes erythrocyte accumulation in an enlarged caudal vein. Moreover, expression of D/A mutations of the putative C-terminal active sites of spp, sppl2 and sppl3 produced phenocopies of the respectiv knockdown phenotypes. Thus our data suggest that all investigated PSHs/SPPLs are members of the novel family of GxGD aspartyl proteases. Furthermore, SPPL2b is shown to be the first member of the SPP/PSH/SPPL family, which is not located within the ER but in endosomal/ lysosomal vesicles.
C: Identification of a Nicastrin homologue
One of the gamma-secretase complex components is the aminopeptidase domain-containing protein Nicastrin. We have now identified a novel member of this protein family related to Nicastrin, which we refer to as Nicalin (Nicastrin-like protein) (Haffner et al., 2004). This 60-kD protein is part of a novel membrane protein complex unrelated to gamma-secretase and binds to pM5, a 130-kD transmembrane protein with no known functional domain. Both proteins are highly conserved in metazoans and plants, localize exclusively to the endoplasmic reticulum, and show almost identical tissue distribution in humans. Functional studies in zebrafish embryos show that Nicalin and pM5 collaborate to inhibit the formation of anterior axial mesendoderm during gastrulation, a process known to be mediated by the Nodal signaling pathway. Inhibition of this pathway by ectopic expression of Lefty could be rescued by the downregulation of pM5. We, therefore, propose the name Nomo (Nodal modulator) for pM5 and suggest that the Nomo/Nicalin complex regulates mesendodermal patterning by antagonizing the Nodal pathway.
Outlook
We are currently investigating the recently published CD147 for its ability to interact and negatively regulate gamma-secretase function. In addition we are studying the physiological function of ß-secretase. The latter may allow us to identify modulators.
Lit.: 1. Aguzzi, A., and Haass, C. (2003). Games played by rogue proteins in prion disorders and Alzheimer's disease. Science 302, 814-818. Capell, A., Steiner, H., Romig, H., Keck, S., Baader, M., Grim, M. G., Baumeister, R., and Haass, C. (2000). Presenilin-1 differentially facilitates endoproteolysis of the b-amyloid precursor protein and Notch. Nat Cell Biol 2, 205-211. Edbauer, D., Winkler, E., Regula, J. T., Pesold, B., Steiner, H., and Haass, C. (2003). Reconstitution of g-secretase activity. Nat Cell Biol 5, 486-488. Geling, A., Steiner, H., Willem, M., Bally-Cuif, L., and Haass, C. (2002). A gamma-secretase inhibitor blocks Notch signaling in vivo and causes a severe neurogenic phenotype in zebrafish. EMBO Rep 3, 688-694. Haass, C. (2004). Take five-BACE and the g-secretase quartet conduct Alzheimer's amyloid b-peptide generation. EMBO J 23, 483-488.Haass, C., Schlossmacher, M. G., Hung, A. Y., Vigo-Pelfrey, C., Mellon, A., Ostaszewski, B. L., Lieberburg, I., Koo, E. H., Schenk, D., Teplow, D. B., and Selkoe, D. J. (1992). Amyloid b-peptide is produced by cultured cells during normal metabolism. Nature 359, 322-325. Haass, C., and Selkoe, D. J. (1993). Cellular processing of beta-amyloid precursor protein and the genesis of amyloid beta-peptide. Cell 75, 1039-1042. Haffner, C., Frauli, M., Topp, S., Irmler, M., Hofmann, K., Regula, J. T., Bally-Cuif, L., and Haass, C. (2004). Nicalin and its binding partner Nomo are novel Nodal signaling antagonists. Embo J 23, 3041-3050. Kaether, C., Capell, A., Edbauer, D., Winkler, E., Novak, B., Steiner, H., and Haass, C. (2004). The presenilin C-terminus is required for ER-retention, nicastrin-binding and gamma-secretase activity. Embo J 23, 4738-4748. Kaether, C., Lammich, S., Edbauer, D., Ertl, M., Rietdorf, J., Capell, A., Steiner, H., and Haass, C. (2002). Presenilin-1 affects trafficking and processing of betaAPP and is targeted in a complex with nicastrin to the plasma membrane. J Cell Biol 158, 551-561.
Lammich, S., Schobel, S., Zimmer, A. K., Lichtenthaler, S. F., and Haass, C. (2004). Expression of the Alzheimer protease BACE1 is suppressed via its 5'-untranslated region. EMBO Rep 5, 620-625. Okochi, M., Steiner, H., Fukumori, A., Tanii, H., Tomita, T., Tanaka, T., Iwatsubo, T., Kudo, T., Takeda, M., and Haass, C. (2002). Presenilins mediate a dual intramembranous gamma-secretase cleavage of Notch-1. Embo J 21, 5408-5416. Shirotani, K., Edbauer, D., Prokop, S., Haass, C., and Steiner, H. (2004). Identification of distinct gamma -secretase complexes with different APH-1 variants. J Biol Chem. Steiner, H., and Haass, C. (2000). Intramembrane proteolysis by presenilins. Nat Rev Mol Cell Biol 1, 217-224. Steiner, H., Kostka, M., Romig, H., Basset, G., Pesold, B., Hardy, J., Capell, A., Meyn, L., Grim, M. G., Baumeister, R., et al. (2000). Glycine 384 is required for presenilin-1 function and is conserved in polytopic bacterial aspartyl proteases. Nat Cell Biol 2, 848-851.